> サイトマップ

本計画班は、構造生物学的なアプローチによって「細胞内タンパク質分解」の分子メカニズムの解明を目指します。周知のように、ユビキチン-プロテアソーム系やオートファジー系における細胞内タンパク質の「分解」は、不要タンパク質の無秩序な破壊ではなく、時間・空間的に厳密にコントロールされた過程です。このような制御は、様々なタンパク質分子の組織的な働きによって実現されていると考えられます。タンパク質分子間の相互作用を解明するためには、立体構造からのアプローチが極めて有効です。
本計画班では、以下の三つのテーマについて研究を進めます。
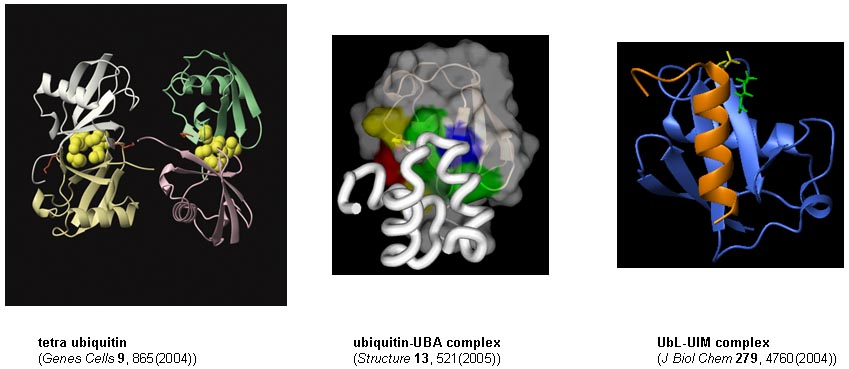
ユビキチン・プロテアソーム系においては、分解されるべきタンパク質にポリユビキチン鎖が付加され、これが「分解シグナル」として働きます。しかし、この「分解シグナル」がどのように認識され、プロテアソームによる「分解」へと到るのか?と言う点に関しては、未だ十分に理解されていません。
我々のグループでは、ユビキチン分子を認識・結合するユビキチン結合ドメインとユビキチンの相互作用について立体構造学的なアプローチで研究を進め、ユビキチンシグナルの認識機構を明らかにすることを目指します。また、ポリユビキチン鎖には重合様式の異なるアイソフォームが存在し、細胞内では各々が異なる意味を持つシグナルとして働くとされています。そこで、アイソフォーム間の立体構造(四次構造)の違いと、ターゲット分子との結合様式の違いについても構造学的な見地から研究を進めます。
[ 上図を拡大する ]
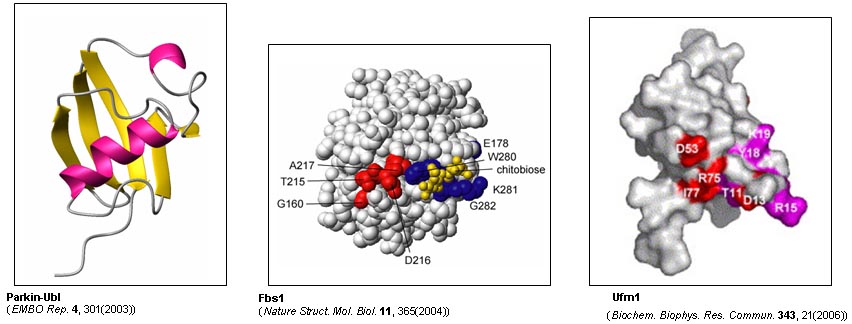
本研究グループは、タンパク質の細胞内分解に関連する翻訳後修飾系(ユビキチン化、NEDD8化、糖鎖修飾など)を対象とする構造生物学的研究を通じて、その作動機構と機能発現の構造的基盤を解明することを目指します。
具体的には、ユビキチンリガーゼ、脱ユビキチン酵素、さらにはNEDD8をはじめとするユビキチン様修飾タンパク質系を対象に構造生物学的研究を行い、構造情報に基づいて、神経変性疾患をはじめとするタンパク質分解系に起因する疾患の発症メカニズムの構造的基盤を明らかにします。また、ユビキチン系を標的とする創薬に向けた基礎的知見を得ることも視野に入れています。
我々のグループは、タンパク質の効率的リフォールディング技術、糖鎖プロファイリングをはじめとする翻訳後修飾の解析技術、糖タンパク質・多サブユニットタンパク質の高度な安定同位体標識技術などの独自の技術を保有しています。さらに、超高磁場NMR装置(920MHz)を利用することにより、natively unfolded proteinや糖鎖修飾タンパク質の構造解析が可能です。
[ 上図を拡大する ]
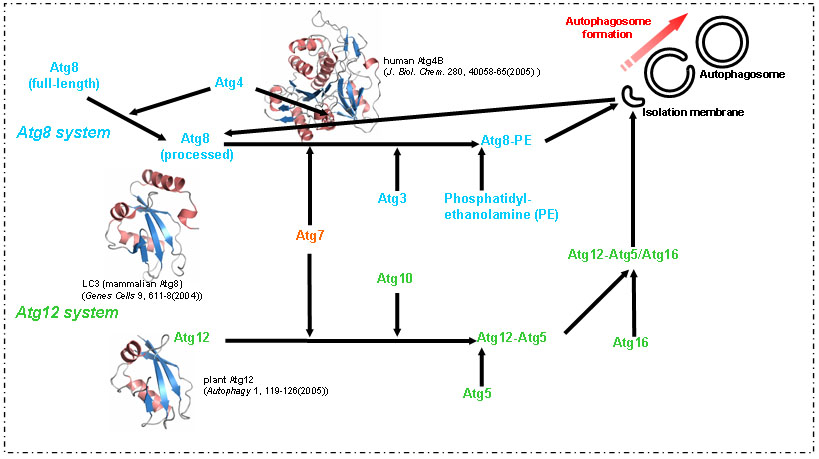
オートファジーの研究は、現・基礎生物学研究所の大隅良典教授らの出芽酵母を用いた先駆的研究によって急速に進歩し、少なくとも16種類のAtg蛋白質群がオートファゴソーム形成に必須であることが見出されました。しかし、これらのAtg蛋白質群がオートファジーの過程において果たす役割およびその機構については、依然として未解明の部分が多いのが現状です。現在までのAtg蛋白質群に関する構造生物学的研究は、もう一つの分解系であるユビキチン-プロテアソーム系に比べて大変遅れており、構造情報が非常に乏しい状態にあります。本研究グループは、Atg蛋白質群を集中的に構造解析し、オートファジーの分子機構解明のための構造的基盤を確立することを目指します。
私たちはこれまでに、二つのユビキチン様修飾系であるAtg8系とAtg12系に関わる8種類のAtg蛋白質のうち、3種類について立体構造を報告してきました。まだ立体構造の報告がない残り5種類のAtg蛋白質について、主にX線結晶構造解析法により構造決定します。そしてこれらAtg蛋白質の溶液中での構造変化や分子間相互作用を、主にNMR法により原子レベルで明らかにします。特に、修飾系の産物であるAtg8- phosphatidylethanolamine結合体、Atg12-Atg5結合体がオートファゴソーム形成に果たす役割を、その立体構造情報から解明することを目指します。
[ 上図を拡大する ]
![]()


